Received 18 June 2004 | Accepted 17 November 2004 | Published 16 December 2004
Reprint
Received 18 June 2004 | Accepted 17 November 2004 | Published 16 December 2004 |
Download Reprint |
Massive increase in the stiffness of the human lens nucleus with age: the basis for presbyopia?
Karl Robert Heys,1,2 Sandra Leigh Cram,3
Roger John Willis Truscott1,2
1Australian Cataract Research Foundation, University of Wollongong, Wollongong, Australia; 2Save Sight Institute, Sydney, Australia; 3BlueScope Steel Institute, University of Wollongong, Wollongong, Australia
Correspondence to: Roger Truscott, Department of Chemistry, Universtity of Wollongong, Wollongong, New South Wales, 2522, Australia; Phone: 61-2-42213503; FAX: 61-2-42214287; email: rjwt@uow.edu.au
Abstract
Purpose: To determine the stiffness of different regions of human lenses as a function of age, and to correlate the biophysical measurements in the lens center with nuclear water content.
Methods: A custom made probe fitted to a dynamic mechanical analyzer was employed to measure stiffness values at 1 mm increments across equatorial sections of individual human lenses. Thermogravimetric analysis was used to determine the percentage water content in the nuclei of human lenses.
Results: There was a pronounced increase in lens stiffness over the age range from 14 to 78. In the nucleus, stiffness values varied almost 1,000 fold over this age range, with the largest change observed in lenses between the ages of 20 to 60. Nuclear stiffness values increased on average by a factor of 450. By contrast, in the cortex the average increase in stiffness was approximately 20 fold over this same time period. In lenses younger than age 30, the nucleus was found to be softer than the cortex. This was true for all six lenses examined. In contrast all lenses older than 30 were characterized by having nuclear values higher than those of the cortex. In lenses over the age of 50, the lens nucleus was typically an order of magnitude more rigid than that of the cortex. The crossover age, when the cortical and nuclear stiffness values were similar, was in the 30s. There was no significant change in the water content of the human lens nucleus from age 13 to age 82.
Conclusions: There is a marked increase in the stiffness of the human lens with age. This is most pronounced in the nucleus. Since in vivo data indicate that the nucleus must change shape significantly during accommodation, it is highly likely that these measured changes in physical properties will markedly diminish the ability of the lens to accommodate, and thus may be a major contributing factor to presbyopia. Since there was no measurable difference in the water contents of the nuclear regions of the lenses, this marked increase in stiffness is not due to compaction of the lens nucleus.
Introduction
The lens is unique in that there appears to be no protein turnover in the center throughout one's lifespan. This lack of turnover in the nucleus leaves it prone to age related changes. Mature lens fibers lack cellular organelles and contain a highly concentrated protein solution to enable light refraction.
Presbyopia, or "old man's eyes," affects almost all people by the age of 50. This disorder involves a loss of accommodative ability, where near focussing becomes impossible. Herman von Helmholtz [1] first expounded the basis for accommodation. In this scenario, contraction of the ciliary muscles leads to a change in the lens curvature. This optical system moves between two states; one where muscular force is applied to the zonules which flattens the lens and allows distant vision, and an accommodated state where zonular tension is released and the lens assumes a more rounded shape to allow focussing on near objects. The exact reason for the development of presbyopia is unknown, and many hypotheses have been proposed. One theory postulates that changes in the ciliary muscle are implicated [2-5] and another school of thought suggests that presbyopia is the result of age related changes to the lens or capsule [6-9]. In the latter hypothesis it is suggested that changes with aging result in the inability of the lens to alter shape from the flattened form, to that of the more rounded accommoded state, when zonular tension is released. Presbyopia has also been described as a geometric disorder resulting from the increased size and volume of the lens and the angle of zonular insertion onto the lens [10]. The vitreous has also been proposed to contribute to accommodation [11].
It has been found that there is an increase in the lens cross sectional area in the accommodated form and that this difference in cross sectional area between accommodative states declines with age [12]. A recent in vitro study found that replacement of a presbyopic lens with a flexible polymer restores accommodative ability [13], strengthening the case that lenticular causes may be a major contributor to presbyopia.
It is now clear that the lens nucleus must change shape significantly to enable transition to the accommodated state [14-16]. The extent of change required by the nucleus is far greater than that of the cortex, which may not change shape to any great extent [16].
Sclerosis, or hardening, of the lens tissue would obviously hinder deformation and would significantly reduce the ability of the lens to undergo the changes required for accommodation. Preliminary data suggests that sclerosis of the human lens center might occur with age [17]. It has been suggested that the changes in physical properties may be due to fiber cell compaction in the nucleus [18], as has been found for several laboratory animal lenses. However, compaction of the lens center does not appear to take place in normal human lenses [19,20] although the number of lenses analyzed in these studies has been small.
A number of methods for analyzing the physical characteristics of lenses have been developed. These include testing using ultrasound wave attenuation [21], and the use of an automated guillotine [22-24]. The main focus of these studies has been the investigation of hardness changes in cataract lenses, since this is an important factor when phacoemulsification is to be employed during surgery. Few studies have looked at how normal lenses change with age. The bulk of studies on normal lenses have used techniques such as applied radial force [8,9,20], or compression/stretching [25-27] tests. These have shown that there is a considerable decline in the ability of the lens to deform with age. However, the use of these techniques is limited due to the inability to differentiate between capsular and lenticular changes, and a limited ability to detect differences throughout the lens. The use of a conical probe to examine changes within the lens showed that an increase in hardness does occur with age [17]. This method involved the use of a miniature dynamometer, in which a probe gradually penetrated the lens. The increasing friction on the probe as it encounters greater depths in the lens is a problem with this method of analysis, and there is also a possible effect of compaction as the probe is inserted deeper into the lens.
The aim of the present study was to generate a cross sectional map of stiffness across the lens and to examine how this profile changes with age. Instrumentation that has been used to analyze materials such as hydrogels was employed. The results of this study are surprising. Stiffness of the lens center was found to increase by approximately 1,000 times and the outer region of the lens by a factor of 50. In addition we found that the nucleus of younger lenses was actually softer than the cortical region. The massive change in the physical properties of the lens center with age was not accompanied by a change in water content.
Methods
Analysis of lens stiffness
Normal lenses covering a broad age range were collected from the Lions NSW Eye Bank at the Sydney Eye Hospital. These were transferred to Wollongong and stored at -80 °C. Lenses (18) between the ages of 14 and 76 years were analyzed with at least 2 lenses per decade. The work was approved by the human research ethics committee at the University of Wollongong.
Lenses were removed from the freezer, partially thawed and then sectioned equatorially. The lens halves were then cored using an 8.5 mm internal diameter trephine incorporating a removable 3.5 mm high steel ring which retained the lens sample (Figure 1 and Figure 2). Once the ring containing the lens was removed from the trephine, if necessary, a razor blade was passed across the top of the 3.5 mm high steel ring to produce a flat surface corresponding to an equatorial section roughly through the middle of each lens. This procedure provided support for the outer edges of the lens sections and also normalized the sizes of all the lens samples.
The lens sample held within the ring was then placed into a sample holder consisting of two pieces of Perspex. The top piece has a central indent in which to position the metal ring (Figure 1D,E), and two metal pins through it allowing it to locate into the base piece. The base piece has a 10x10 1 mm grid pattern drilled in it. This allowed movement of the sample holder by 1 mm. The bottom of the base piece of the sample holder was shaped to fit the stage of the penetration clamp of the dynamic mechanical analyzer (DMA).
The instrument used for the analysis was a Q800 DMA (TA Instruments, New Castle, DE). A custom made steel cylindrical probe was designed with a flat bottom and a diameter of 0.4 mm. This small probe enabled measurements to be taken every millimeter across the surface of the lens samples. All readings were performed at 22 °C.
Stiffness measurements on the lens sections were performed using a controlled force test. The probe was positioned on the lens surface and the force was ramped from 0 to a maximum force of 3 mN, at a rate of 1 mN per minute, while the change in displacement over the course of the run was monitored. Typically the displacement was of the order of 750 μm. These measurements commenced at the edge of the sample ring and then moved by 1 mm increments across the lens surface. Three runs across each sample were taken wherever possible. This consisted of a run across the center of the sample and then subsequent runs parallel to, but 1 mm above and below the initial traverse. It did not affect the results significantly if measurements were commenced in the lens center or the edge (data not shown). The nuclear value used for calculations of shear modulus was either the fourth or the fifth measure of stiffness across the lens, whichever was greater.
Each measurement took approximately 3 min to complete, so three runs of eight measurements involved a total analysis time of 90 min per lens. To alleviate any possible sample dehydration, a plastic housing containing a large piece of moistened foam rubber was used to surround the probe.
Analysis of water content
Measurements of lens nuclear water content were performed using thermo-gravimetric analysis (TGA) at the University of Technology, Sydney. The instrument used was an SDT 2960 (TA Instruments).
Whole lenses were partially thawed in a petri dish, and then cored using a 4 mm trephine. The core had the top and bottom ends (approximately 1 mm) removed leaving a cylinder 4 mm in diameter and approximately 4 mm in height. This material was transferred to a platinum crucible, and immediately placed in the TGA for analysis.
TGA analysis involved heating from ambient temperature to 300 °C at 5 °C per minute. The loss in mass of the sample was monitored over time, and the total water loss was determined at the point where the derivative value of loss in mass dropped to a value near to zero. Typically this was at about 190 °C. The remaining mass was considered to represent protein.
Results
This study was undertaken with a view to determining the stiffness profile across human lenses and to examine if there were any consistent alterations with age. A DMA was chosen as the instrument of choice. Since the cross sectional area of human lenses is quite small, a probe with a narrow (0.4 mm diameter) bore was designed and manufactured. A special base plate was also designed to enable measurements at 1 mm intervals across each lens, and a trephine constructed as shown in Figure 1 to produce an equatorial section of tissue constrained within an 8.5 mm metal ring that was independent of the original size of the lens. This apparatus allowed the measurement of 8 readings across each lens (Figure 2). In most cases three passes were made across the diameter of the lens sections and the results at each point were averaged. In order to measure the stiffness at each point, a protocol was adopted in which the displacement of the probe was determined following ramping of the force from 0 to 3 mN at 1 mN per min.
The results were converted into values of shear modulus using the equation:
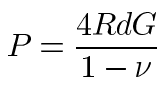
where "P" is total load (N), "R" is radius of indentor (m), "d" is maximal depth of penetration (m), "G" is shear modulus (Pa), and "ν" is Poisson's ratio for the material [28]. For an elastic rubber like material, we assumed that ν=0.5 and that we were using a perfectly rigid flat punch in frictionless contact with a semi-infinite (large enough that edge effects are not encountered), fully elastic material.
The results of a typical analysis of a 64 year old lens are shown in Figure 3. It is clear that there is a substantial increase in the stiffness of the lens as the probe is moved closer to the lens center. As would be expected, the profile is roughly symmetrical about the central point.
In contrast the results of an analysis of a 19 year old lens are shown in Figure 4. The values for shear modulus were lower in this case and the central region of the lens was less stiff than the cortex. This was a surprising finding. Again the profile was symmetrical about the central point. Since these two lenses gave stiffness profiles that were quite different in appearance, we applied this DMA technique across an age range from 14 to 76 years. A summary of the results is depicted in Figure 5. Because the changes in lens stiffness were of such magnitude, it was only possible to compare them using log scales. These illustrate clearly that there is a consistent age dependent increase in stiffness in both the cortex and the nucleus of human lenses.
The magnitude of these changes in the two regions was, however, quite different. In the cortex there was, on average, an approximately 20 fold increase in lens stiffness over the age range from 14 to 76 (Figure 5 and Table 1). The lowest cortical value (48.5 Pa) was determined in a 14 year old lens and the highest (2,577 Pa) in a 76 year old lens. This corresponds to a 53 fold change. In the nucleus this increase in lens stiffness was much more pronounced, amounting to, on average, 450 fold in the space of just over 50 years (Figure 5). The values ranged from the softest nuclear value (25.7 Pa) measured in a 20 year old lens, to the hardest (23,954 Pa) in a 73 year old lens, a 930 fold increase.
The nuclear values of lens stiffness in younger lenses were consistently lower than those of the cortex. For the age group younger than 30 years, of which there were 6 individuals, the mean shear modulus value for the nucleus was 39.0±13.8 Pa, compared to 98.3±64.5 Pa for the cortex. Analysis of these results with a paired Student's t-test showed that this was statistically significant (p<0.05). In lenses older than 60 years, the mean value for the nucleus was 17,400±4,900 Pa, and 2,040±710 Pa for the cortex. Analysis of these results with a paired Student's t-test showed that this was also statistically significant (p<0.02). Since the nuclear values increase at a more rapid rate than those of the cortex, there must be a crossover point. As is illustrated in Figure 5, this crossover occurred at approximately age 30-35. Although more lenses need to be analyzed to confirm this trend, the largest changes seem to be observed in lenses between the ages of 20 and 60.
In order to confirm that the shear modulus values measured with the custom made probe were correct, the shear modulus values of a hydrogel of similar stiffness were also determined. The elastic modulus (E) of the dimethylacrylamide based hydrogel had previously been measured by compression testing on the DMA and was found to be 20±5 kPa. The hydrogel was tested using the custom made indentation probe in the same manner as the lenses, and a shear modulus of 6.29 kPa was determined. When this was converted to elastic modulus by the standard expression
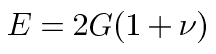
giving an elastic modulus value of 18.9 kPa. This is in very good agreement with the results found by compression testing.
These are remarkable changes in physical properties of the human lens. The reasons for these are not yet known, but some features can be ruled out. There was, for example, no evidence for lens compaction. If lens compaction were taking place with age, the percentage of protein in the nucleus should increase. As shown in Figure 6, no significant change in the percentage of protein was noted over the whole age range. It should be noted that the lenses used for analysis of stiffness were, in several cases, the contralateral lenses of those used for the determination of nuclear hydration. These are indicated on the graph (Figure 6).
The finding of no apparent change in percentage protein with age in the human lens is in agreement with that of van Heyningen [19]. Others have found variable results on water content with age [29,30], in this regard human lenses are clearly different from those of common experimental animals. Rats [19], rabbits [19,20], and cattle [19] all show a clear increase in lens protein content as the animals age.
Discussion
In this study large changes in the biophysical properties of normal human lenses as a function of age have been demonstrated. The observed increase in stiffness was much more evident in the center of the lens.
Although nuclear compaction does not appear to be associated with the huge changes in stiffness that we documented, there is not yet a clear picture of the molecular changes that are responsible. It is clear, however, that there is a marked parallel between the findings of increased lens stiffness described here, and the well known age dependent alterations in accommodation [1,3,4,26,31]. This is illustrated in Figure 5 and Figure 7. In addition, the maximal change in lens power clearly shows a marked change from teenage years, such that by age 60 human lenses show essentially no change in focal length with stretching [26]. As has been summarized in recent articles by Croft, Glasser and Kaufman [31,32], the basis for presbyopia remains unclear but has been largely ascribed to two possible causes involving either the ciliary muscle or the lens. Other theories have also been advanced. These include a geometric theory based on the age dependent alteration in the sites of attachment of the zonules [10,33] coupled with lens growth and multifactorial [34] postulates. The vitreous has also been suggested to play a part in facilitating asymmetric alterations in lens dimensions and forward movement of the lens during accommodation [11]. It is well established that the consistency of the vitreous alters with age [35].
It is clear that our results support changes to the properties of the lens as being implicated. As documented by Koretz and coworkers [16] and others [14,15] normal accommodation in humans involves changes to the shape of the lens nucleus. If the nucleus becomes progressively stiffer with age, it is not surprising that the lens becomes much more difficult to deform. This is especially true if the change in stiffness involves nearly 3 orders of magnitude.
Of considerable interest is the fact that these results reveal changes in lens hardness starting from the teenage years and continuing into old age. These are most dramatic in the nucleus. The shortest attainable focal length of the human lens increases almost linearly as a function of age [31]. The recent in vitro demonstration that accommodation can be restored to human lenses by implanting intraocular lenses made from a flexible polymer [13] also supports a key role for the lens in presbyopia.
Quite remarkably, DMA analysis of the younger lenses showed that the nuclear region was softer than the cortex in lenses younger than age 30. This is in contrast to those lenses aged over 30. This finding is in agreement with those of Fisher [9] but at variance with those of Pau and Kranz [17], although these authors did not show separate data for cortex and nucleus.
It should be noted that while this study was underway, Weeber and Eckert in the Netherlands were conducting a similar investigation also using DMA analysis [36]. Although their methodology was slightly different from ours, the results that they obtained seem to mirror those obtained here. For example, they reported, using a smaller number of lenses, an approximately 10,000 fold increase in shear modulus in the nucleus over a similar age range, and in their abstract describe that the nuclear values for stiffness in young lenses were less than the cortical values.
All lenses used in this study had been stored frozen. It is possible that fresh lenses may behave differently and this will be investigated in a future study. Analysis of fresh lenses is more difficult technically due to their "stickiness" making coring and lens sectioning more difficult.
While our results demonstrating increased hardness of isolated human lenses mirror those found by others for decreased accommodative power measured in the eyes of volunteers of different ages, several caveats should be noted. First, the process of accommodation involves stretching, rather than the compressive forces that were used for DMA analysis. Second, we cannot discount a contribution from proteolysis. Proteases may become active after we section the lenses prior to DMA measurements, although it would be expected that levels of endogenous proteases in the nucleus will be low, especially in older lenses. In addition as noted above, freezing may affect the results and our calculations of lens stiffness in the nuclei of the youngest lenses may be an overestimate due to the limitations of the current methodology.
It is conceivable that the physical changes we have measured in these lenses are part of a spectrum of age related ocular alterations, all of which may contribute to presbyopia. The huge increases in stiffness could also be the result of some other factors that over time may affect lens biochemical processes. For example, lens transport [37,38] could be influenced by the regular lens deformation associated with accommodation and, if this process is compromised could, in turn, affect the biophysical properties of the lens.
It is not understood what factors determine lens stiffness and why these should change with age. In very young lenses this could be a reflection of the initial difference in protein composition in the nucleus compared with the cortex [39,40]. For example, γ-crystallins are concentrated in the nucleus since they are predominately synthesized prenatally. It is clear however that both cortex and nucleus show similar alterations over time, although the magnitudes are quite different.
A quite unexpected finding was that the nuclei of young lenses (prior to age 30), were less stiff than those of the cortices. Given the requirement for substantial nuclear deformation during accommodation, one is tempted to suggest that such a difference may be present to facilitate the process of lenticular shape change.
Protein oxidation can also be ruled out as the reason for the large increase in human lens hardness with age. Analysis of protein sulfhydryl (data not shown, [41]) and protein methionine sulfoxide contents [42] have revealed no significant oxidation of human lens proteins past age 60. Once again, laboratory animals such as rodents appear to be poor models for aging of the human lens since in these animals there is evidence of considerable protein oxidation with age [43,44].
Progressive losses in soluble α-crystallin with age [45,46] could be implicated in the physical changes noted, since this protein is a molecular chaperone and has been proposed to play an important role in maintaining optical clarity by binding to, and sequestering, other lens proteins as they denature over time. This may also contribute to the activities of other key proteins, for example channel proteins such as connexins and aquaporin 0. Since protein aggregation within the cell may also lead to changes in the relative amounts of free and bound water, this factor may also be implicated. Age dependent changes in the composition of fiber cell membranes [47] could also influence lens stiffness. These factors are to be investigated in the next phase of this study. This work is to be repeated with a higher sampling frequency so as to correlate the size of the zone of stiffness with the barrier to diffusion that also develops at middle age. This barrier appears responsible for the later onset of age related nuclear cataract [48].
Acknowledgements
We wish to acknowledge Professor Geoff Spinks and Professor Hugh Brown from the Engineering Department at the University of Wollongong for their generous help with the DMA work, Mr. Raj Devasahayam from Sydney Lions Eye bank for providing the lenses, and Steve Cooper from the Science Faculty workshop for making the lens cutter and sample holders. Roger Truscott is an NHMRC Senior Research fellow. Karl Heys is supported by a scholarship provided by the Save Sight Institute. The DMA was purchased via a grant from the Australian Research Council (LIEF, Hugh Brown). This work was supported by grants from the NIH R01 EY013570 and NHMRC 307615.
References
1. von Helmholtz, H. Uber die akkomodation des auges. Albrecht von Graefe's Arch Ophth 1855:1-74.
2. Duane A. Are the current theories of accommodation correct? Am J Ophthalmol 1925; 8:196-202.
3. Fincham EF. The mechanism of accommodation. Br J Ophthalmol 1937; 8(Suppl):5-80.
4. Fincham EF, Walton J. The reciprocal actions of accommodation and
convergence. J Physiol 1957; 137:488-508. ![]()
5. Eskridge JB. Review of ciliary muscle effort in presbyopia. Am J
Optom Physiol Opt 1984; 61:133-8. ![]()
6. Hess C. Observations concerning accommodation organs. Klin Monatsbl Augenheilkd 1904; 42:309-15.
7. Gullstrand A. Mechanism of accommodation. In: Southall JPC, editor. Helmholtz's treatise on physiological optics. Vol 1: appendices. Rochester (NY): Optical Society of America; 1924.
8. Fisher RF. The significance of the shape of the lens and capsular
energy changes in accommodation. J Physiol 1969; 201:21-47.
![]()
9. Fisher RF. The elastic constants of the human lens. J Physiol
1971; 212:147-80. ![]()
10. Koretz JF, Handelman GH. How the human eye focuses. Sci Am 1988;
259:92-9. ![]()
11. Coleman DJ. Unified model for accommodative mechanism. Am J
Ophthalmol 1970; 69:1063-79. ![]()
12. Strenk SA, Strenk LM, Semmlow JL, DeMarco JK. Magnetic resonance
imaging study of the effects of age and accommodation on the human lens
cross-sectional area. Invest Ophthalmol Vis Sci 2004; 45:539-45.
![]()
13. Koopmans SA, Terwee T, Barkhof J, Haitjema HJ, Kooijman AC.
Polymer refilling of presbyopic human lenses in vitro restores the
ability to undergo accommodative changes. Invest Ophthalmol Vis Sci
2003; 44:250-7. ![]()
14. Brown NAP, Bron AJ. Lens disorders: a clinical manual of cataract diagnosis. Oxford: Butterworth-Heinemann; 1996.
15. Dubbelman M, Van der Heijde GL, Weeber HA, Vrensen GF. Changes in
the internal structure of the human crystalline lens with age and
accommodation. Vision Res 2003; 43:2363-75. ![]()
16. Koretz JF, Cook CA, Kaufman PL. Accommodation and presbyopia in
the human eye. Changes in the anterior segment and crystalline lens with
focus. Invest Ophthalmol Vis Sci 1997; 38:569-78. ![]()
17. Pau H, Kranz J. The increasing sclerosis of the human lens with
age and its relevance to accommodation and presbyopia. Graefes Arch Clin
Exp Ophthalmol 1991; 229:294-6. ![]()
18. Al-Ghoul KJ, Nordgren RK, Kuszak AJ, Freel CD, Costello MJ,
Kuszak JR. Structural evidence of human nuclear fiber compaction as a
function of ageing and cataractogenesis. Exp Eye Res 2001; 72:199-214.
![]()
19. Van Heyningen R. The human lens. 3. Some observations on the
post-mortem lens. Exp Eye Res 1972; 13:155-60. ![]()
20. Fisher RF, Pettet BE. Presbyopia and the water content of the
human crystalline lens. J Physiol 1973; 234:443-7. ![]()
21. Tabandeh H, Wilkins M, Thompson G, Nassiri D, Karim A. Hardness
and ultrasonic characteristics of the human crystalline lens. J Cataract
Refract Surg 2000; 26:838-41. ![]()
22. Tabandeh H, Thompson GM, Heyworth P. Lens hardness in mature
cataracts. Eye 1994; 8:453-5. ![]()
23. Tabandeh H, Thompson GM, Heyworth P, Dorey S, Woods AJ, Lynch D.
Water content, lens hardness and cataract appearance. Eye 1994; 8:125-9.
![]()
24. Heyworth P, Thompson GM, Tabandeh H, McGuigan S. The relationship
between clinical classification of cataract and lens hardness. Eye 1993;
7:726-30. ![]()
25. Pierscionek BK. In vitro alteration of human lens curvatures by
radial stretching. Exp Eye Res 1993; 57:629-35. ![]()
26. Glasser A, Campbell MC. Presbyopia and the optical changes in the
human crystalline lens with age. Vision Res 1998; 38:209-29.
![]()
27. van Alphen GW, Graebel WP. Elasticity of tissues involved in
accommodation. Vision Res 1991; 31:1417-38. ![]()
28. Sneddon IN. The relaxation between load and penetration in the axisymmetric boussinesq problem for a punch of arbitrary profile. International Journal of Engineering Science 1965; 3:45-57.
29. Siebinga I, Vrensen GF, De Mul FF, Greve J. Age-related changes
in local water and protein content of human eye lenses measured by Raman
microspectroscopy. Exp Eye Res 1991; 53:233-9. ![]()
30. Lebensohn JE. Biochemistry of the lens. Arch Ophthal 1936; 15:216-21.
31. Croft MA, Glasser A, Kaufman PL. Accommodation and presbyopia.
Int Ophthalmol Clin 2001; 41:33-46. ![]()
32. Glasser A, Croft MA, Kaufman PL. Aging of the human crystalline
lens and presbyopia. Int Ophthalmol Clin 2001; 41:1-15.
![]()
33. Farnsworth PN, Shyne SE. Anterior zonular shifts with age. Exp
Eye Res 1979; 28:291-7. ![]()
34. Weale R. Presbyopia toward the end of the 20th century. Surv
Ophthalmol 1989; 34:15-30. ![]()
35. Balazs EA, Denlinger JL. Aging changes in the vitreous. In: Sekuler R, Kline D, Dismukes K, editors. Aging and human visual function. New York: Liss; 1982. p.45-57.
36. Weeber HA, Eckert G. Stiffness distribution within the human crystalline lens and its function with age. ARVO Annual Meetng; 2004 Apr 25-29; Fort Lauderdale, FL.
37. Mathias RT, Rae JL. The lens: local transport and global
transparency. Exp Eye Res 2004; 78:689-98. ![]()
38. Sweeney MH, Garland DL, Truscott RJ. Movement of cysteine in
intact monkey lenses: the major site of entry is the germinative region.
Exp Eye Res 2003; 77:245-51. ![]()
39. Thomson JA, Augusteyn RC. Ontogeny of human lens crystallins. Exp
Eye Res 1985; 40:393-410. ![]()
40. Bours J, Wegener A, Hofmann D, Fodisch HJ, Hockwin O. Protein
profiles of microsections of the fetal and adult human lens during
development and ageing. Mech Ageing Dev 1990; 54:13-27.
![]()
41. Truscott RJ, Augusteyn RC. The state of sulphydryl groups in
normal and cataractous human lenses. Exp Eye Res 1977; 25:139-48.
![]()
42. Sochaski MA, Jenkins AJ, Lyons TJ, Thorpe SR, Baynes JW. Isotope
dilution gas chromatography/mass spectrometry method for the
determination of methionine sulfoxide in protein. Anal Chem 2001;
73:4662-7. ![]()
43. Askren CC, Yu NT, Kuck JF Jr. Variation of the concentration of
sulfhydryl along the visual axis of aging lenses by laser Raman optical
dissection technique. Exp Eye Res 1979; 29:647-54. ![]()
44. Kuck JF, Yu NT, Askren CC. Total sulfhydryl by raman spectroscopy
in the intact lens of several species: variations in the nucleus and
along the optical axis during aging. Exp Eye Res 1982; 34:23-37.
![]()
45. McFall-Ngai MJ, Ding LL, Takemoto LJ, Horwitz J. Spatial and
temporal mapping of the age-related changes in human lens crystallins.
Exp Eye Res 1985; 41:745-58. ![]()
46. Roy D, Spector A. Absence of low-molecular-weight alpha
crystallin in nuclear region of old human lenses. Proc Natl Acad Sci U S
A 1976; 73:3484-7. ![]()
47. Borchman D, Byrdwell WC, Yappert MC. Regional and age-dependent
differences in the phospholipid composition of human lens membranes.
Invest Ophthalmol Vis Sci 1994; 35:3938-42. ![]()
48. Truscott RJ. Age-related nuclear cataract: a lens transport
problem. Ophthalmic Res 2000; 32:185-94. ![]()
49. Bruckner R, Batschelet E, Hugenschmidt F. The Basel longitudinal
study on aging (1955-1978). Ophthalmo-gerontological research results.
Doc Ophthalmol 1986; 64:235-310. ![]()
50. Duane AJ. Normal values of the accommodation of all ages. J Am Med Assoc. 1912; 59:1010-3.
51. Donders FC. On the anomalies of accommodation and refraction of the eye; with a preliminary essay on physiological dioptrics. London: New Sydenham Society; 1864.
52. Hamasaki D, Ong J, Marg E. The amplitude of accommodation in
presbyopia. Am J Optom Arch Am Acad Optom 1956; 33:3-14.
![]()
53. Pierscionek BK, Weale RA. Presbyopia - a maverick of human aging.
Arch Gerontol Geriatr 1995; 20:229-40. ![]()